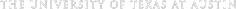One of the most fossil-rich and well known sites in paleoanthropology, the Hadar Research Area is located ~300 km northeast of Addis Ababa, Ethiopia. The name Hadar derives from the nearby Kada Hadar tributary that feed into the Awash River1,2. A thin greenbelt surrounding the Awash River is the only greenery among Hadar’s badlands of rolling hills and occasional desert scrub brush. Few plants can survive Hadar’s arid conditions, where temperatures routinely climb as high as 110ºF (40º C) during the day1.
Hadar sits in the southwest of the Afar Triangle, an ~200,000 km2 geologically active area where several tectonic plates are rifting apart (i.e. Nubian, Arabian, and Somalian plates)3-5. Many of Hadar’s discoveries have been the result of this triple rifting system. As the tectonic plates jostle against one another, a low lying basin has formed at the junction of the Ethiopian Plateau on the west and the Somalian Plateau to the south/southeast2,5. Sediments from all over the area gradually fall and accumulate within the basin. The Awash River Valley, situated in this basin, is a rich source of late Miocene to the early Pleistocene2 sediments that were mostly transported by streams2,6. Numerous preserved volcanic tuffs, evidence of past tectonic activity, provide good source material for remarkably precise dating of the Hadar fossils1,5.
French Geologist Maurice Taieb first discovered Hadar in 19682 as part of his work on the geological evolution of the Awash River Valley4. A few years later, Taieb struck up a conversation with a young doctoral candidate about Taieb’s work in Ethiopia. This graduate student, named Donald Johanson, was intrigued by Taeib’s fossil discoveries and the 2collaborated on survey work in 19724. Realizing Hadar’s fossil potential, Taieb joined with Johanson and French paleontologist Yves Coopens to organize the International Afar Research Expedition, and the team of French and American scientists began working at Hadar in the fall of 19731,4,7.
The team began to recover hominin fossils almost immediately, including some of the earliest evidence of hominin bipedalism known at that time4. In 1973, Johanson identified a 3.4 million year old (Ma) fossilized knee (AL 129) that exhibited bipedal adapatations4. Other finds included the famous “Lucy” skeleton, which was over 40% complete; and a large cache of early hominin individuals known as the First Family. By 1977, over 250 hominin specimens had been recovered8, and explorations at Hadar were suspended to allow for proper description of the Hadar material. The team’s analyses concluded that the Hadar (and Laetoli) remains were from a previously unknown hominin that possibly represented the oldest yet known human ancestor4,9. The fossils were assigned to a new species called Australopithecus afarensis10.
Excavations resumed at Hadar in 1990 and continue today under the Hadar Research Project8,11. To date, nearly 400 hominin specimens have been collected at Hadar, and all but one have been identified as Au. afarensis. The 1994 discovery of a fragmentary palate dating to 2.3 Ma, tentatively identified at Homo habilis12, may place one of the earliest known Homo species at Hadar. Crude stone flakes discovered in association with the Homo specimen also suggest that Hadar may be home to some of the earliest evidence for hominin stone tool use13.
Over 400 collection sites have been established since the first fossils were found at Hadar in the late 1960, and more than 90% of all Au. afarensis material comes from the Hadar area14. The importance of Hadar in the study of human evolution continues to grow as more scientific explorations take place. The 2 fossil hominins represented at Hadar may lie at the heart of major radiation events in human evolutions, including the proliferation of Au. afarensis and the first appearance of Homo. Future discoveries at Hadar, therefore, are sure to increase our understanding of human origins.
Hadar has been a fossils hot spot since anthropologists first identified a hominin fossil at the site in 1973. In fact, the geologic record of Hadar may hold the keys to some of most fascinating periods in human evolution. Fossils derived from Hadar span between 3.4 and 2.36,13,15 million years ago (Ma), and may document the existence of at least 2 extinct hominins that lie at the core of the modern human lineage – Australopithecus afarensis and possibly early Homo.
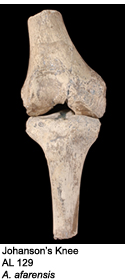 Throughout the mid to late 1970s, the International Afar Research Expedition (IARE) discovered a large number of Au. afarensis fossils at Hadar. The species was first discovered at Laetoli in 1934, but wasn’t properly identified as Au. afarensis until the Hadar excavations. The first hominin specimen was discovered by Donald Johanson during an afternoon survey in October 1973. Johonson identified a proximal tibia and a distal femur sticking out of the ground. To his surprise, the two ends articulated together perfectly, indicating that they were from the same individual4. To his even greater surprise, the femur appeared to angle inwards toward the body’s midline, a clear indication that a bicondylar angle was present4. The bicondylar angle is a distinct carrying angle between the hip and the knee that places the foot under the body’s center of gravity during the walking cycle16. The articulated knee joint found by Johanson, known as A.L. 129 or “Johanson’s Knee”, dates to ~3.4 Ma and helped to push back the estimated evolution of bipedalism, from 1.8 Ma to over 3 Ma1,18.
Throughout the mid to late 1970s, the International Afar Research Expedition (IARE) discovered a large number of Au. afarensis fossils at Hadar. The species was first discovered at Laetoli in 1934, but wasn’t properly identified as Au. afarensis until the Hadar excavations. The first hominin specimen was discovered by Donald Johanson during an afternoon survey in October 1973. Johonson identified a proximal tibia and a distal femur sticking out of the ground. To his surprise, the two ends articulated together perfectly, indicating that they were from the same individual4. To his even greater surprise, the femur appeared to angle inwards toward the body’s midline, a clear indication that a bicondylar angle was present4. The bicondylar angle is a distinct carrying angle between the hip and the knee that places the foot under the body’s center of gravity during the walking cycle16. The articulated knee joint found by Johanson, known as A.L. 129 or “Johanson’s Knee”, dates to ~3.4 Ma and helped to push back the estimated evolution of bipedalism, from 1.8 Ma to over 3 Ma1,18.
During the next field season, Donald Johanson and Tom Gray were surveying and mapping possible fossil localities at Hadar once again. The two were just about to return to camp, when Johanson suggested they explore a nearby gully. The dry stream bed was empty of fossils, but a bone lying partway up the slope of the gully caught Johanson’s attention4. The bone was part of a hominin arm. A few feet away, they discovered fragments of a skull, femur, vertebrae, and pelvis. They marked the site as A.L. 288.
After three weeks of excavations, the A.L. 288 specimen was reassembled at camp. Over 40% of the skeleton was recovered, with nearly every bone in the body represented1. The team quickly realized they discovered one of the most unique and complete hominin specimens ever found. Preliminary analysis of the pelvis and femur suggested that the skeleton was that of a small sized adult female, about 107 cm (3’6”) tall. The skeleton was officially named A.L. 288-1, after the fossil locality where it was found, but she quickly picked up the nickname “Lucy”, after the Beatles’ song, “Lucy in the Sky with Diamonds,” which was playing at camp the night she was found1. She is also known as “Dinquinesh”, which means “wonderful onel” in the local Amharic language of Ethiopia14. Today, Lucy is permanently housed at The Ethiopian National Museum in Addis Ababa, Ethiopia.
“Lucy” and “Johanson’s Knee” provided some of the first hominin postcranial material over 3 Ma, and for the first time, anthropologists could reconstruct the arms and legs of early hominins. The femora of both A.L. 129 and Lucy exhibited a bicondylar angle4, and Lucy’s nearly complete left os coxae (hip bone) and sacrum appeared similar in shape to that of modern humans. However, Lucy’s upper limbs retained features typically seen in arboreal primates such as chimpanzees, including a cranially oriented scapula (shoulder blade), curved phalanges, and relatively long arms (i.e., although Lucy’s arm and leg length ratio is intermediate between modern humans and chimpanzees)6,16,17. Lucy, therefore, suggested that Au. afarensis possessed a mosaic of derived and primitive traits. Nevertheless, the derived condition of Lucy’s pelvis and limbs strongly confirmed the bipedal nature of Au. afarensis, and provided some of the first conclusive evidence that small bodied, small brained (~434 cc) hominins walked bipedally at least 3.2 Ma4. (For more information, visit the Who is Lucy and Step by Step: Evolution of Bipedalism lessons).
IARE hypotheses about Au. afarensis were further substantiated with the discovery of the A.L. 333 cache, a collection of Au. afarensis fossils nicknamed “The First Family” that dates to ~3.2 Ma. In the fall of 1975, IARE returned to Hadar accompanied by National Geographic photographer David Brill, a french husband and wife filmmaker team, and several anthropology students4. In early November 1975, Johanson led the team to a small gully marked as A.L. 333 where two hominin teeth had been found the previous day by Mike Bush. While Brill set up his camera equipment, one of the filmmakers pointed out a calcaneous and femur resting on the suface, halfway up the the hillside4. It turned out that the gully and surrounding slopes were littered with hominin fossils4,9. To date, more than 250 Au. afarensis skeletal elements have been recovered from the A.L. 333 locality, representing at least 9 adults, 3 adolescents, and 5 juveniles (the youngest approximately 2 years old)6,14,19,20.
Accumulation of such a large hominin assemblage within a single geologic horizon continues to pose interesting questions. Early analysis suggested that the hominins had fallen victim to a catostraophic flash flood19. Recent studies focusing on the A.L. 333 deposits, however, argue for a low-energy, shallow flooding event21. In addition, a number of the fossils exhibit signs of weathering, and/or irregular fractures typical of surface exposure. It is likely, therefore, that the hominins died at the site from an unknown cause, were exposed to the surface for relatively short period, followed by burial in slow moving fluvial sediments22. Savenging of the material is likely, as the frequency of articulated specimens is quite low, and the majority of the assemblage consists of limb and cranial elements suggesting a felid predator22. The unique assemblage continues to draw scientific interest, and even produce hominin fossils 35 years after its initial discovery.
In 1992, the first relatively complete skull of an Au. afarensis was discovered at Hadar by Yoel Rak14. A.L. 444 is one of the most complete and largest Au. afarensis cranium found to date, probably belonging to male with an estimated brain size of 530 cc and dating to ~3 Ma14,23. A fragmented male mandible, frontal bone, and partial upper limb skeletons that include a nearly complete ulna was discovered at A.L. 438, and dates to ~3 Ma23. The Hadar collection also include female cranial specimens, including the nearly complete skull A.L. 822 found in 2002 (~3.1 Ma)24, and a nearly intact female face consisting of a fragmented mandible and cranial base known as A.L. 417 (~3.25 Ma)23. Among the post cranial specimens are a humerus dating to ~3.4 Ma (A.L. 137)9, and a left proximal femur from A.L. 1287.
To date, about 400 hominin specimens have been recovered at Hadar, providing an invaluable resource for understanding early hominins. The diverse Hadar collection can be used to determine intraspecies variation, sexual dimorphism, diet, and social structures. The collection may also indicate a period of evolutionary status8. Over the 400,000 years the species occupied Hadar, Au. afarensis retained both derived features associated with bipedalism, and primitive traits such as small brain size (415 cc) and powerful upper body6. The combination of these traits suggests that Au. afarensis engaged in terrestrial bipedalism and some degree of arboreality. Whatever future explorations at Hadar may find, there is no doubt that the evidence found at Hadar shook loose previously held hypotheses about hominin evolution, including the timing of hominin bipedalism. Whereas scientists once believed large brains were the hallmark of human ancestry, Lucy and other Au. afarensis fossils successfully proffered bipedalism as the oldest shared trait in the human lineage.
Homo sp. (Possibly Homo habilis)
On November 2, 1994, two members of the Hadar Research Project named Ali Yesef and Maumin Allahendu discovered bone and stone flakes at the bottom of gully at Hadar locality A.L. 66614,13. Not far from the A.L. 444 locality, A.L. 666 is located within the the Makaamitalu drainage basin near a Awash River tributary. Deposits at this site date to at least ~2.33 Ma, based on 40Ar/39Ar methodology25. The A.L. 666 site constitutes the youngest occurance of hominins at Hadar.
 Among the bone fragments discovered by the researchers were two maxillary pieces broken along the intermaxillary suture14. Thus, the two havles, when articulated together, comprise a complete maxilla. When discovered, the maxilla had been exposed to the surface only recently, and was relatively well preserved13. Further excavation recovered isolated teeth and tooth fragments that, upon examination, could be easily rearticulated into the maxilla13. In total, most of the A.L. 666 teeth were present or recovered, including both canines, and all cheek teeth. Fully erupted third molars and visible wear patterns on the teeth indicate A.L. 666 was a mature adult, while the relatively large size of the maxilla, sinus cavity, and associated dentition indicates that the individual was probably male13.
Among the bone fragments discovered by the researchers were two maxillary pieces broken along the intermaxillary suture14. Thus, the two havles, when articulated together, comprise a complete maxilla. When discovered, the maxilla had been exposed to the surface only recently, and was relatively well preserved13. Further excavation recovered isolated teeth and tooth fragments that, upon examination, could be easily rearticulated into the maxilla13. In total, most of the A.L. 666 teeth were present or recovered, including both canines, and all cheek teeth. Fully erupted third molars and visible wear patterns on the teeth indicate A.L. 666 was a mature adult, while the relatively large size of the maxilla, sinus cavity, and associated dentition indicates that the individual was probably male13.
Initially, A.L. 666 was believed to be Au. afarensis, based largely on the fossil’s antiquity and discovery location. In a 1997 analysis, A.L. 666 was cautiously assigned to the species Homo habilis25, a designation that continues to promote discussion. Many researchers prefer to assign A.L. 666 to an unidentified species of Homo.
This contentious assignment of A.L. 666 to H. habilis is based largely on apomorphies common to all Homo. For example, unlike the rectangular-shaped maxilla and tapered midface of australopiths, the A.L. 666 midface is relatively square, and the dental arcade more parabolic13. Likewise, australopith palatal length, width, and depth are long, narrow, and shallow (respectively), but A.L. 666 exhibits a relatively long, wide, and deep palate13,25. Other derived features include moderate subnasal prognathism, slightly flatter incisors, relatively narrow first molars and rhomdoial second molars, and thinner molar enamel25. While these traits are typical of all Homo, A.L. 666 tends to fall outside the range of variation for most species, to the exception of H. habilis. In comparision with other H. habilis specimens, A.L. 666 appears most similar to the adult male KNM-ER 180525. If the taxonomic assignement of H. habilis stands on this premise, then perhaps the differences between the A.L. 666 maxilla and other H. habilis specimens can be attributed to sexual dimorphism25. The 1997 analysis goes on to argue that the taxonomic assignment to H. habilis is the least problematic based on the currently known comparative record25. The recovery of more Homo specimens of the same geological age as A.L. 666 are needed to better understand the variation between the A.L. 666 specimen and other hominins.
The only Homo fossil found thus far at Hadar, A.L. 666 remains one of the youngest hominin specimens to derive from one of the most fossil rich sites in all of Africa. At 2.3 Ma, it may not only be one of the earliest known specimens of Homo, but also one of the oldest hominins associated with manufactured lithics. Other stone tools have been discovered at Gona and Bouri (in 1992 and 1997 respectively), but the A.L. 666 specimen constitutes the oldest hominin to be confidently associated with manufactured lithics and faunal assemblage13.
No artifacts have been associated with A. afarensis at Hadar.
The archaeological record at Hadar begins at locality AL 666, which dates to 2.33 Ma. Approximately 20 lithics, consisting of flakes and bifacial choppers were discovered on the surface in close association with a fragmented hominin mandible in 1994. The surface flakes were remarkably well-preserved and still had sharp edges, indicating recent surface exposure.
In total, 34 artifacts were found at the AL 666 locality, including 14 lithics recovered from the Kada Hadar member, the uppermost strata of the Hadar geologic formation. All the artifacts (as well as the hominin maxilla) were found in close proximity to one another, clustered within a 90 x 80 cm area in a layer no more than 9 cm thick.
Many of the AL 666 artifacts exhibit simple striking platforms and preserved cortical material, all fetures consistent with Oldowan tools, the earliest form of stone tool technology. While the assemblage included at least 3 bi-facial choppers, most of the material constituted flakes typically used for food processing. However, the flakes show no evidence of use or retouching. The lithics were most likely manufactured from rounded chert and basalt river cobbles. In at least 1 instance, excavators rearticulated a flake onto its original core.
Even though the Hadar flakes appeared unused, the AL 666 assemblage also contained several large mammalian bones, which suggest the site might have been used for butchering. In addition, small striae found on a bovid scapula have been interpreted as cut marks made when the muscle was cut from the bone by a stone tool.
The lithic and mammalian assemblage at Hadar indicates that stone tools were manufactured at Hadar by at least 2.3 Ma. The 2.6 Ma stone tools discovered at nearby Gona currently represent the oldest known tool assemblages. The AL 666 lithics, however, represents the oldest associated between hominin and faunal remains. It’s not possible to determine if maxilla found at the site represents an individual who actually produced the stone tools, but the presences of these lithics does imply that hominins had a precision grip by 2.3 Ma. It also implies some form of culture, since stone tool production was probably a learned behavior.

The Hadar hominins lived in a taxonomically rich habitat influenced by oscillations between wet and dry environmental episodes between 4-2 Ma. Along with the early hominins, Hadar’s Plio-Pleistocene inhabitants thrived in a variety of ecological zones that included tropical forest, wetlands, and dry grasslands. A few species are present throughout the entire Plio-Pleistocene, while others species appear only during excessive wet or dry periods. Tracking changes in the frequency of stenotopic species can assist in a holistic reconstruction of ancient habitats, including speciation events and climatic fluctuations either on a local or regional level. Folivores and frugavores were attracted to the open woodlands, bushlands and wooded grasslands present during the relatively low seasonality wet periods that existed approximately 3.42 – 3.26 Ma. Animals whose diets consisted of rich grasses, leaves and branches include various antelope species (e.g., impalas, dwarf antelope, duikers, and extinct forms similar to modern wildebeests and waterbucks), giraffes, gazelles, and the extinct three-toed horse. Large mammals include extinct elephant species, black and white rhinos, and several extinct forms of suids (i.e., pigs). Present during periods of flooding were aquatic animals such mollusks, crabs, and cane rats that live in thick reeds near standing water, along with crocodiles, otters, and hippopos typically found in deep fresh water. Carnivores such as hyenas, jackals, and predatory cats (e.g., servals and an extinct form similar to the saber-toothed cat) likely moved into the area following their prey. Among the many birds, reptiles and rodents, also present were dietary specialists such as the root-eating porcupines, and fruit-eating primates such as the colobus monkey, extinct baboons, and A. afarensis.
Between 3.2 and 2.9 Ma, Hadar’s climate was relatively dry, initiating an increase in arid-adapted populations that roamed open grasslands, such the antelope, gazelle, buffalo, bison, and other forms of wild cattle. The dry conditions also opened up new ecological niches for species not previously seen at Hadar, such as burrowing animals like rabbits, hares, badgers, mongoose, and an extinct rodent similar to the modern mole rat. In addition to the hyenas and saber-toothed cats seen in previous depositional periods, new predators like the panther were introduced to Hadar during this period. Just prior to 2.9 Ma, colobus monkeys and baboons return to Hadar.
Comparing the frequency, or change in frequencies, of species present at certain times can help to identify climate shifts. For example, a decrease in the abundance of arboreal feeders, such as the colubus monkey and Parapapio (an extinct baboon species), coupled with an increase in terrestrial grazer populations between 4 and 2.9 Ma may signal a general shift from wet wooded areas to drier, more open habitats such as grasslands. Indeed, by ~3 Ma species typical of wooded habitats with low seasonality (e.g, impalas and colobus monkeys) are almost completely absent from the fossil record, as are wetland fauna such as waterbucks (Reduncins). Meanwhile, populations of arid-adapted grazers and mixed feeders such as antelopes and wildebeests (Alcelaphini and Antelopini respectively) increase substantially over the same period.
Despite these environmental changes, several eurytopic species are occupied Hadar during all depositional periods, including an extinct African elephant (Elephas recki), the carnivorous striped hyenas (Crocuta dietrichi), and the mixed feeder giraffe (Sivatherium marusium). Among those species present prior to 2.9 Ma are several wild pig species (Kolpochoerus afarensis and Notochoerus euilus), wild cattle (Ugandax coryndoni), the three-toed horse (Eurygnathohippus hasumense), an extinct large-bodied baboon-like primate (Theropithecus darti), and A. afarensis.
Approximately 3.42 Ma, A. afarensis encompasses over 30% of the total large mammalian faunal sample and then drops in frequency to approximately 15% by 3.3 Ma. The A. afarensis population continues to decrease, making up only about 5% of the faunal sample (based on minimum number of individuals) between 3.26 and 3.2 Ma. From the time of Lucy at 3.2 Ma, until the species disappears from Hadar around 2.9 Ma, the A. afarensis population comprises less than 10% of faunal assemblage. The consistent presence of A. afarensis fossils in deposits indicating all types of habitats suggests that the species was eurytopic, meaning that A. afarensis was adapted to a wide variety of environmental conditions. Faunal and paleoecological evidence suggests that the A. afarensis population drops during wet periods such as the SH depositional period as a result of flooding, and that population increases may related to a drying-out period, such as during the Denen Dora period. Scientists have not been able to ascertain a clear reason for the disappearance of A. afarensis at Hadar after 2.9 Ma due to gaps in the geologic record.
Nearly all mammalian species present at Hadar prior to 2.9 are replaced by other species, possibly a result of migrations, extinctions, or speciation events trigged by severe climatic or geological shifts that effected food supplies (i.e. changes in vegetation). Hadar began to ardify around 2.9 Ma, and by 2.0 Ma more arid-adapted and fresh-grass grazers were present at Hadar than at any other time. A high percentage of edaphic grass feeders migrated into Hadar for the first time, including antelopes, camels, and several extinct species of gazelles and wild cattle. If the AL 666 maxilla does represent early Homo, then Homo habilis appears at Hadar for the first time beginning around 2.3 Ma.
The Hadar site is named for deposits exposed along the Kada Hadar and Ounda Hadar tributaries of the Awash River. These deposits are notable for the Pliocene fossil record contained within them. The site is located in the Afar depression, an area where three rifts come together: the East African Rift, the Red Sea Rift, and the Gulf of Aden rift. As a result of this junction, there is considerable tectonic and volcanic activity in the area.
The geological formation at Hadar, represented in the diagram on the left, is comprised mainly of silty clays and sands, deposited by rivers and, when a lake was present, along shores and deltas. The formation is divided into four members. Over the Basal Member sits the volcanic Sidi Hakoma Tuff, dated to 3.40±0.03 Mya. The Sidi Hakoma Member lies above the Sidi Hakoma Tuff and ends at Triple Tuff 4 (3.22±0.01 Mya). A layer of Kadada Mournou Basalts (3.28±0.04 Mya) gives further resolution to the Sidi Hakoma member, which contains the hominin localities A.L. 137 and A.L. 417. Next, the Denen Dora Member lies between Triple Tuff 4 and the Kada Hadar Tuff (3.18±0.01 Mya). The Denen Dora Member contains hominin locality A.L. 333. Finally, the Kada Hadar Member is the best studied section of the Hadar Formation. It contains Bouroukie Tuff 2 (2.92±0.03 Mya) and is topped by Bouroukie Tuff 3 (2.33±0.07 Mya).
The Kada Hadar Member contains notable hominin fossil localities A.L. 288, A.L. 438, A.L. 444, and A.L. 666 (with associated artifacts). During the period when this member was deposited, a lake expanded across the Hadar study area a total of three separate times. These events are partially responsible for the sandy fluvial deposits in the Kadar Hadar Member. The flooding and receding cycles of this lake also affected the environment in which hominins such as A. afarensis were living.

Between 3.45 and 2.9 Ma, Hadar’s environment remained a mosaic of open and closed woodland and bushland habitats, with various expansions and regressions of dry open grasslands and wet closed forests (i.e., woodlands). On at least three occasions, a large lake expanded (i.e. transgressed) across the area, leaving behind mollusks and reed pollen in the fossil record. The changing habitats influenced by climatic and geologic conditions, also attracted a variety of animals that migrated in and out of the area as environmental conditions changed, contributing to the subsistence of several stenotopic species.
Hadar’s current vegetation consists of plants adapted to hot, arid environments such as acacia trees and desert scrub brush, but 4 – 3.5 Ma, Hadar was covered in open and closed wooded areas, including a large open flat floodplain covered with lush grasses and an expansive lake surrounded by a riparian forest. The currently badlands topography results from an ancient river system that sustained a wider range of floral and faunal species than those that currently exist at Hadar. The trees and shrubs found in Hadar’s paleoenvironment were adapted for warm, wet conditions, while the grasses and underground storage organs (i.e., roots and tubers) present during later depositional periods, around 2.9 Ma, were adapted for more arid and hotter climates. For the majority of the Hadar formation, most of the grasses were probably sustained from water sources such as lakes and wetlands. After 2.9 Ma, the previously low occurrence of C4 grasses.
During most of the Miocene (23.8 – 5.3 Ma), much of Africa was covered in thick tropical forests, but at the Miocene-Pliocene boundary around 5 Ma, the general climate of the earth was moving toward a cooling event. In Africa, this meant a reduction in heavily wooded areas, such as tropical forests, and the spread of more open woodlands, savannas, and grasslands. In the early Pliocene, Hadar was still relatively closed and wet, meaning that the environment consisted of heavily wooded areas and received large amounts of rainfall each year, probably resulting from strong monoons that form regularly off the east coast of Africa each year. In fact, between 3.42 and 3.35 Ma, Hadar received a higher than average amount of rainfall resulting in less seasonal weather and the expansion of wet habitats such as humid woodlands, wetlands, and lake margins.
During the mid Pliocene, between 3.4 and 3.22 Ma, Hadar’s habitat was a mosaic of mostly dry and riparian woodlands, shrublands, and a few open grasslands. At some point during this period, between 3.35 and 3.26 Ma, the wet climate aided in the development of a major wetland event, including at least two major lake transgressions and regression in the Hadar floodplain. Paleolake Hadar formed amidst a dry open woodland and savanna habitat, and its marshy borders were full of reeds, sedges, and mollusks.
Hadar’s climate continued to fluctuate between wet and arid periods, until the end of the Denen Dora depositional period (~3.2 Ma) when Hadar’s environment began a general drying trend, resulting in more seasonality (i.e., a rainy season), and more dry open woodlands and wooded grasslands. By 3 Ma, Hadar’s once tropical rainfall was reduced to only 31 – 35 inches (80-90 cm) per annum, whereas today’s average tropical rainforest receives approximately 100 inches of rain per year. Between 2.9 and 2.33 Ma, Hadar appeared as a flat open plain covered with savanna grasslands with small patches of trees around small isolated water sources. Fresh grasses and USO were the major sources of vegetation attracting many arid-adapted grazing animals such as antelopes and other bovines, which in turn attracted large numbers of carnivores.
After the final regression of Paleolake Hadar, the major water source became the Awash River system, and riverine forest and wetlands became more prominent by 3.22 – 3.18 Ma. The A. afarensis population increases dramatically during this time, including the famous “Lucy” individual. Animals adapted for wetland environments disappear from the fossils record shortly after 2.9 Ma, indicating that Hadar’s climate became increasingly arid. By the time Hadar entered another wet period resulting in a large wetland across the area, the fauna common during previous depositional periods was nearly complete replaced by new species.
Climate changes and major shifts in habitats may have influenced migration patterns, as evidenced by a near complete absence in Busidima fossil record of large bodied mammals common in the Hadar Formation (> 2.9 Ma) within the Busidima fossil record (<2.35 Ma), including the replacement of A. afarensis with early Homo. No A. afarensis fossils dating younger than 2.5 Ma have been found at Hadar. Prior to 2.5 Ma, A. afarensis is found in all depositional period between the upper Basal and the upper Kada Hadar members, indicating that A. afarensis was euytopic rather than a habitat specialist, an advantageous characteristic at Hadar given the climatic fluctuations.
While other animals affected by environmental changes would migrate in and out of the area, A. afarensis could stay in one place and exploit the changing habitat. In fact, A. afarensis fossil have been identified within a variety of ecological niches, from open wet woodlands to relatively drier open shrublands. On the other hand, the early Homo specimen exisited during a time of open grassland, when Hadar was known to be more arid. However, the exact association between the disappearance of A. afarensis, the advent of Homo, and Hadar’s changing habitats remains unknown as a large section of sediments are missing from the geologic record that could otherwise help reconstruct Hadar’s habitat during that time (2.9 – 2.35 Ma).